Summary
One major premise of landscape ecology is that the natural environment tends to be heterogeneous i.e., patchy. Consequently, studies have concentrated on the distribution of various trees in various locations. In Australia, researchers have sought to understand the distribution of mistletoes and other trees in large areas of natural vegetation (Calder, 1997; Reid, 1990; Turner, 1991). They have also established that the relationship between mistletoes and host plants in Australia presents two unique kinds of interactions. First, mistletoe is a hemiparasite plant that gets its water and nutrients from the host plant. Second, there is a mutualistic relationship between mistletoe and birds, specifically the Mistletoebird. This bird disperses mistletoe seeds. On the other hand, Mistletoebird feeds on the fleshy fruits of the plant. This creates mutual benefits for the two species. In addition, Mistletoe plants offer shelter and food to other species of birds in Australian forests and woodlands.
This study seeks to test patchiness by investigating the distribution of trees and mistletoes in a large area of natural vegetation in Australia. Since mistletoes are hemiparasite plants that feed on shrubs and trees, the study will not only examine the distribution of species, but also an ecological relationship between parasites and hosts.
Objectives
The main objective of the study is to investigate the distribution of the Drooping Mistletoe Amyema pendulum across an urban bushland landscape in relation to variation in vegetation type and the abundance of host tree species. In this regard, the study will address the following questions:
- Does the density of trees (stems) differ between vegetation types in the study landscape?
- Does the proportion of trees with mistletoe infestation differ between vegetation types?
- What is the spatial pattern of mistletoe parasitism at the level of individual trees?
Study Area
The study was conducted at the Royal Botanic Gardens, Cranbourne, a large area of heathland and heathy woodland at Cranbourne. The location has served as a critical place for many ecological studies in the past. Such studies have included a floristic vegetation study, studies of the ecology of Swamp Rats and Southern Brown Bandicoots, resource partitioning by small mammals, the foraging ecology of New Holland Honeyeaters, and studies of frogs.
Methods
Healthy Woodland (drier vegetation on sandy areas) and Wet Heath (lower-lying areas, drainage lines) areas were chosen for the study. These areas are represented in the Garden brochure. The major tree species found in the heathland include Coast Manna Gum E. pryoriana and Mealy Stringybark E. cephalocarpa.
Stratification of transects
A snapshot survey (a single field trip) was used to collect all data for the study. Transects were stratified across the landscape to sample Heathy Woodland and Wet Heath vegetation types. Every group collected data from at least one allocated transect. They aimed to sample several transects in every type of vegetation. These samples were used for data analysis and reporting.
Care was taken not to damage the vegetation in the shrubland.
Data Collection
The group located a suitable transact position within the allocated area and marked a 100M transact by using a tape measure. Data were collected within a range of 20 m wide on both sides of the track. This area represented a plot size of 0.4 ha. The group members systematically walked along the transect line, checked and counted all stems of trees (in cases where trees had more than a single trunk). Data recorded included the following:
- the total number of tree stems within the transect area
- for every stem that had mistletoes present, details recorded were:
- the available number of mistletoe plants
- whether the mistletoe plants were alive or dead
- the level of branch on which the mistletoe plant was located (primary, secondary, tertiary)
The main species of mistletoe common in the bushland was the Drooping Mistletoe Amyema pendulum. The group engaged in a closer observation in order to identify the differences in the foliage of the mistletoe and eucalypts. In addition, researchers also focused on identifying any dead trees, what killed them and under what circumstances. They noted how mistletoe was attached to the eucalypt branch. It was imperative to note all variations in the study area based on topography, soils, the density of trees, moisture availability and vegetation types.
Data analysis
Data were analyzed with SPSS and Excel.
Results
Differences in the density of stems
The mean (average) density of stems for each transect.
The mean score for dry heath was 67.25 while the mean score for wet heath was 123.70.
Independent-samples t-test.
An independent-samples t-test was conducted to compare scores for dry heath and wet heath. There was a significant difference in scores for dry heath and wet heath with the p-value of 0.015, which was less than 0.05.
Does the proportion of trees with mistletoe infestation differ between vegetation types?
The proportion of trees (stems) in each transect that had mistletoe present.
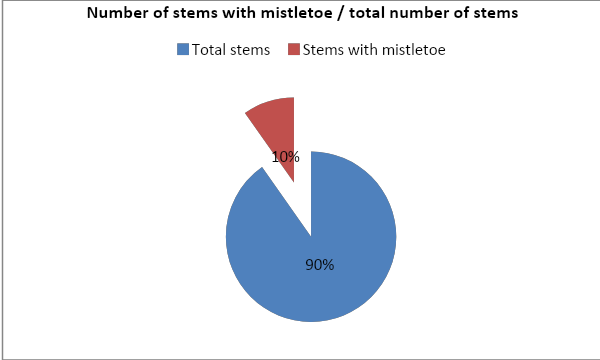
The proportion of trees (stems) in each transect that had mistletoe present was 10% (221) relative to the total number of stems 90% (2044).
An independent-samples t-test was conducted to compare scores for mistletoe in dry heath and wet heath. There was no significant difference in scores for dry heath and wet heath with the p-value of 0.29, which was higher than 0.05.
What is the spatial pattern of mistletoe parasitism at the level of individual trees?
The proportion of mistletoes, which were alive vs. dead.
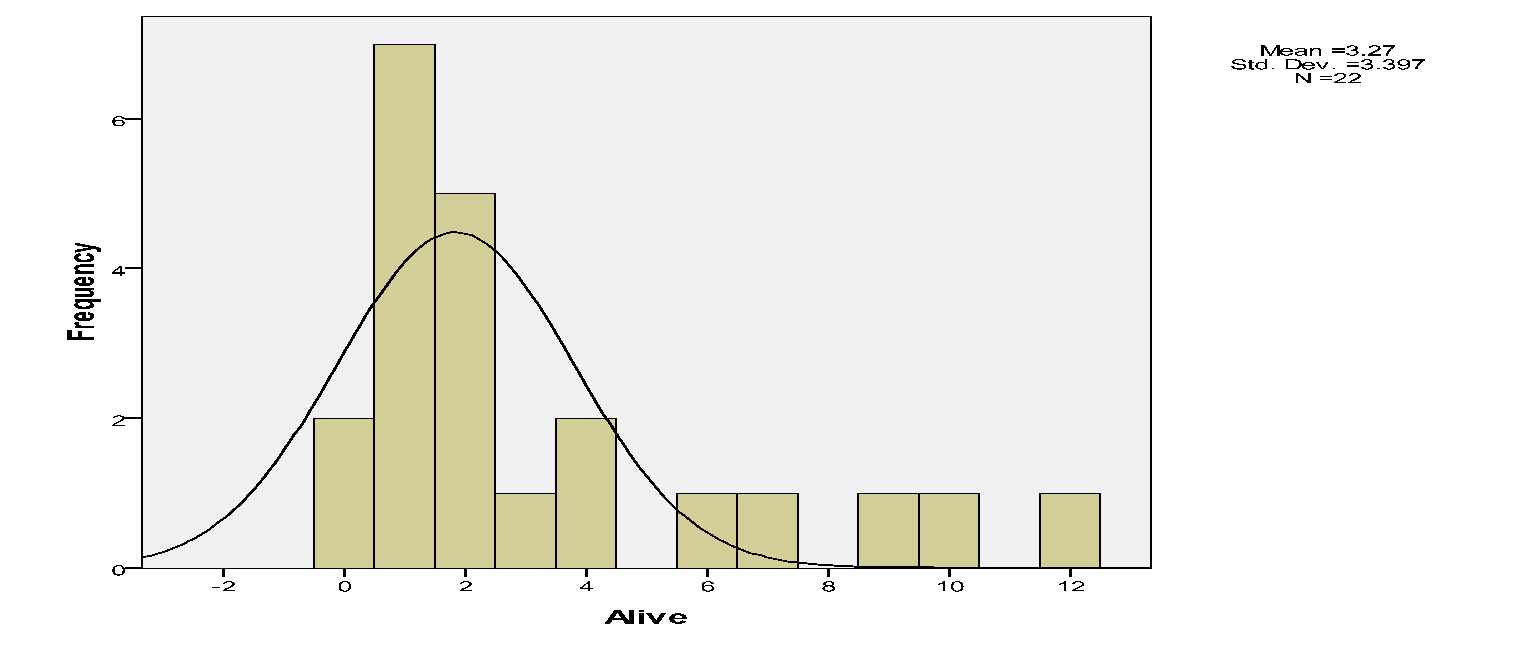
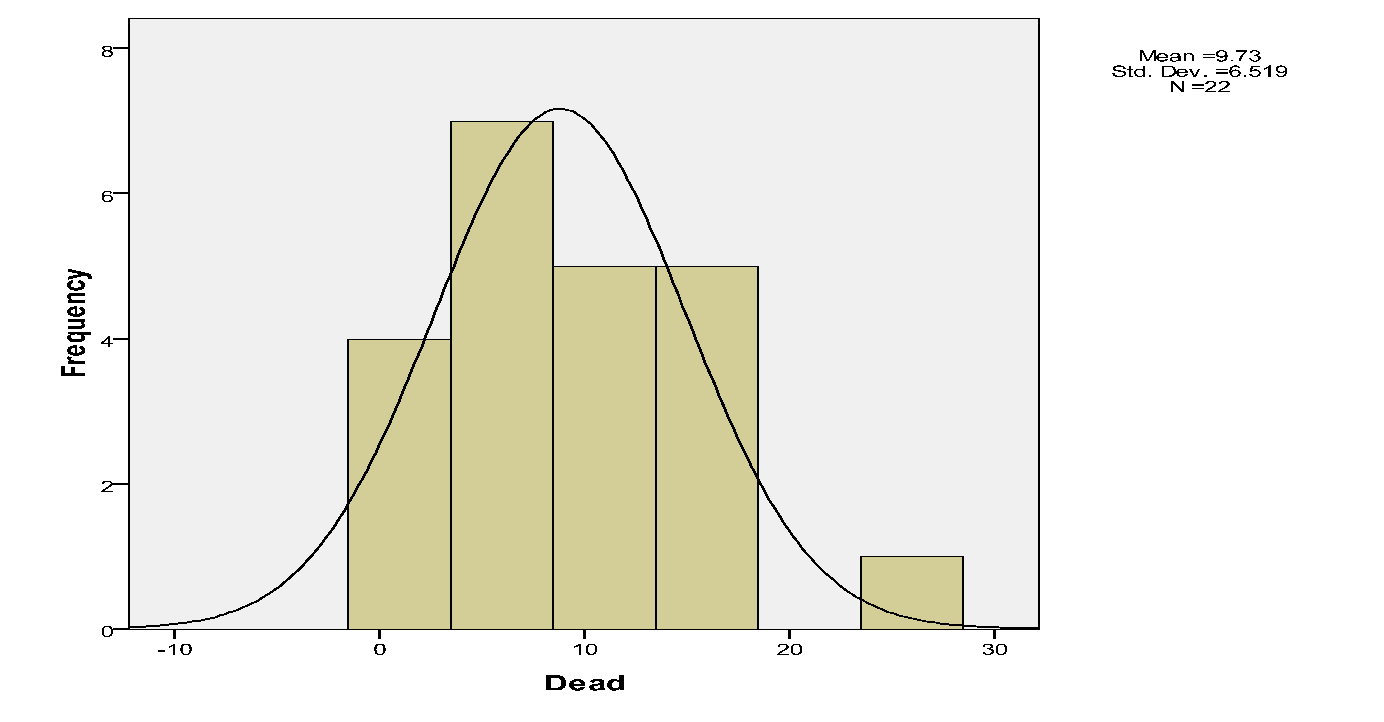
There were few live mistletoes (72) relative dead ones (214).
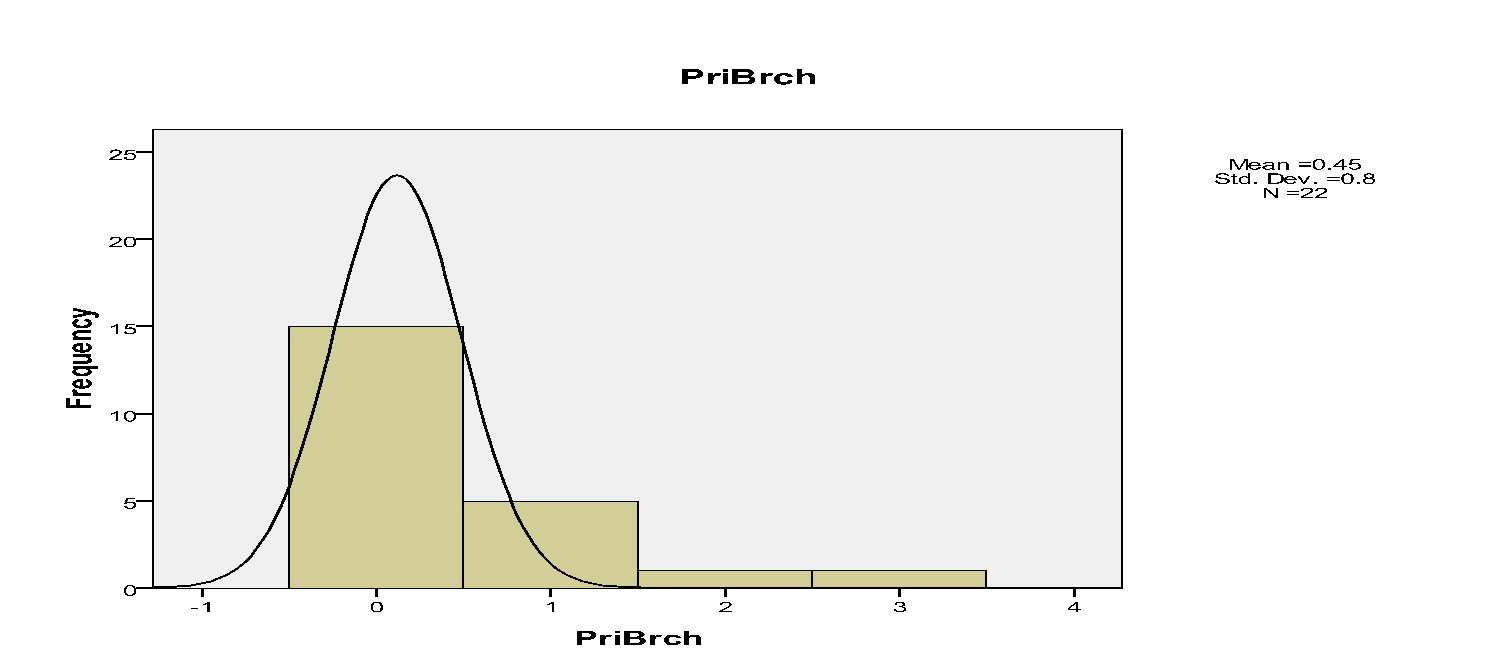
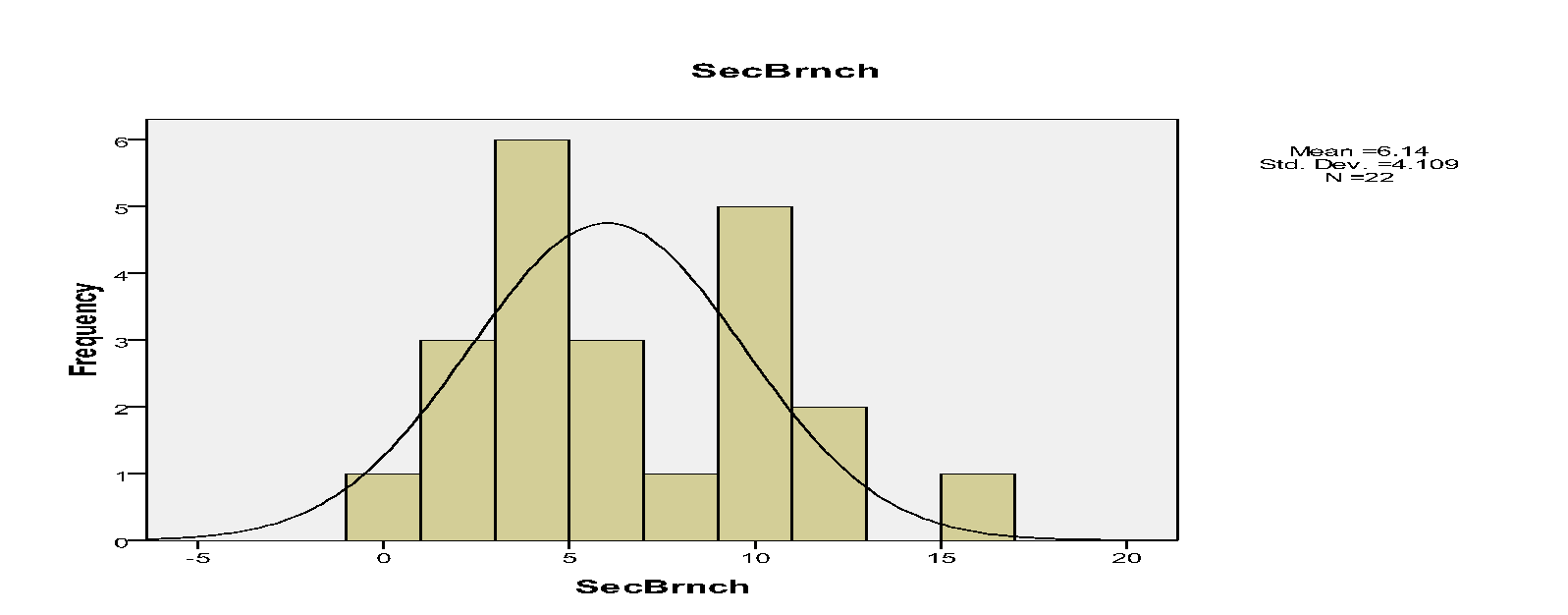
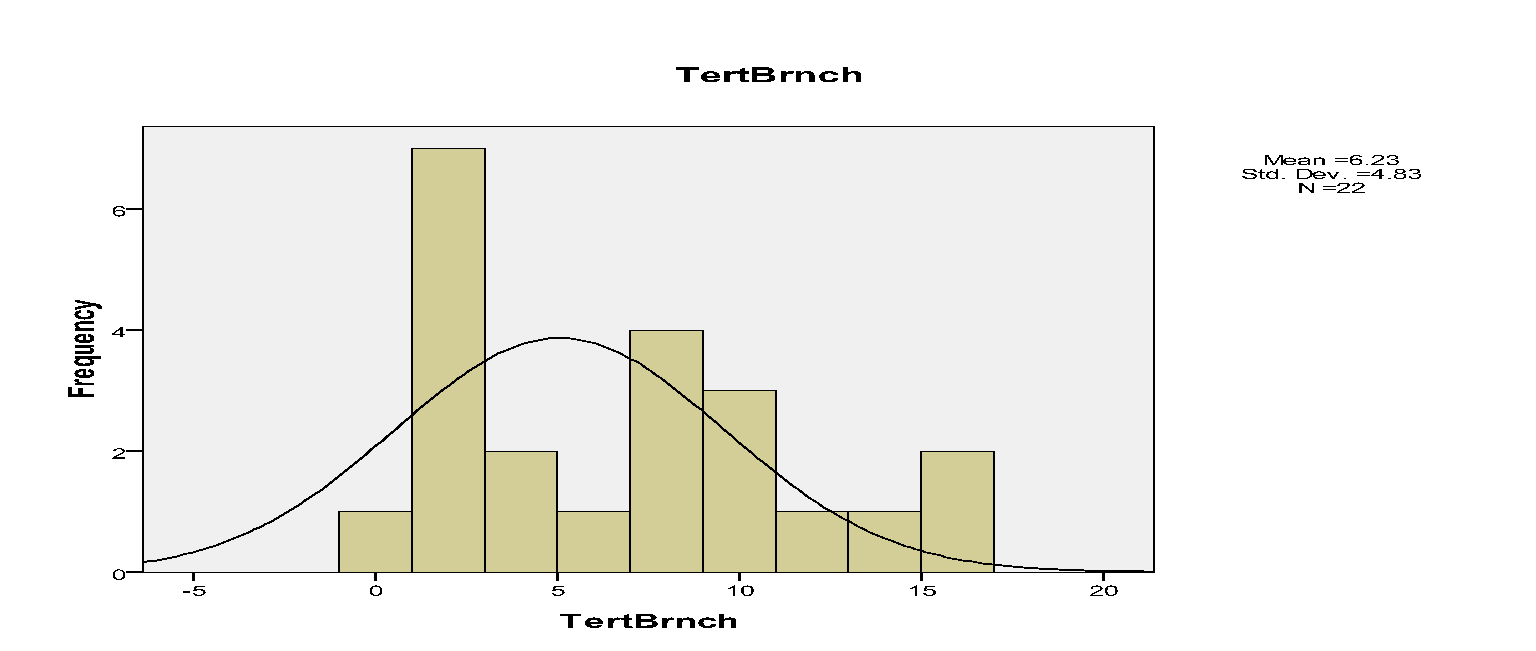
The histograms indicated that spatial occurrences of mistletoes varied to the branches within trees. For instance, mistletoe plants, which were recorded on primary, secondary and tertiary branches, were 10, 135 and 137 respectively.
Discussion
This study confirmed the underlying premise that natural environments are heterogeneous (i.e. patchy) in landscape ecology. Hence, we accepted the study hypothesis that distributions of mistletoes were heterogeneous or patchy and not evenly distributed. An independent-samples t-test was conducted and confirmed a statistically significant difference (p-value of 0.015) between the distribution of stems in dry heath and wet heath. Therefore, the density of trees or stems differed significantly between vegetation types in the study landscape. Several studies have supported such patchy distributions in mistletoes (Press and Phoenix, 2005; Turner, 1991; (Jurskis, Turner and Jurskis, 2005).
The proportion of trees with the mistletoe infestation differed between vegetation types as the study results indicated. For instance, the proportion of trees (stems) in each transect that had mistletoe present was 10% (221) relative to the total number of stems 90% (2044). These figures further support previous study outcomes by other researchers. A further independent-samples t-test was conducted to compare scores for mistletoe in dry heath and wet heath. The results indicated that there were no significant differences in scores for dry heath and wet heath with the p-value of 0.29. Hence, various vegetation types had different mistletoe infestations.
The spatial pattern of mistletoe parasitism at the level of individual trees indicated marked differences. For instance, the primary branches had 10, secondary branches had 135 while tertiary branches had 137 mistletoes.
Large numbers of mistletoes were observed in the wet heath than in the dry heath. This suggests that abiotic factors, such as soils, water and light and biotic factors (dispersal vector) have critical influences on the distribution of the Drooping Mistletoe across the landscape. Press and Phoenix noted that mistletoes had influences on abiotic environments in which they were found. They acted as ecosystem engineers by causing significant changes in both biotic and abiotic elements because they acted as nesting places for birds and other animals. In this regard, Watson observed that mistletoes were critical resources in forests and woodlands where they were found (Watson, 2001).
In addition, few Drooping Mistletoes were located on the primary branches relative to secondary and tertiary branches. Hence, Drooping Mistletoes thrived better in some positions than others. Secondary and tertiary branches on trees tend not to be disturbed. Jurskis, Turner and Jurskis suggested that mistletoes increased in such undisturbed locations (Jurskis et al, 2005). Therefore, such branches offered better habitats for mistletoes.
In conclusion, the study supported previous findings that mistletoes had heterogeneous or patchy distributions in places they were found. In addition, their distributions varied across dry heath and wet heath, as well as in the primary, secondary and tertiary branches of plants. They influenced abiotic factors and were important resources in forests and woodlands (Press and Phoenix, 2005).
Reference List
Calder, M 1997, ‘The mistletoe plant’, Victorian Naturalist, vol. 114, pp. 96-97.
Jurskis, V, Turner, RJ and Jurskis, D 2005, ‘Mistletoes increasing in ‘undisturbed’ forest: a symptom of forest decline caused by the unnatural exclusion of fire?’, Australian Forestry, vol. 68, no. 3, pp. 226-235.
Press, M and Phoenix, G 2005, ‘Impacts of parasitic plants on natural communities’, New Phytologist, vol. 166, no. 3, pp. 737–751. Web.
Reid, N 1990, ‘Mutualistic interdependence between mistletoes (Amyema quandong) and spiny-cheeked honeyeaters and mistletoebirds in an arid woodland’, Australian Journal of Ecology, vol. 15, pp. 175-90.
Turner, R 1991, ‘Mistletoe in eucalypt forest – a resource for birds’,. Aust. Forestry, vol. 54, pp. 226-235.
Watson, D. M. 2001, ‘Mistletoe – a keystone resource in forests and woodlands worldwide’, Annual Review of Ecology and Systematics, vol. 32, pp. 219-249.
Map of the study area