There is increasing evidence that bacteria can “talk” to each other, and in gram-negative bacteria, the “language” used for monitoring and responding to their own abundance has recently been termed quorum sensing. ( Fuqua, 269-75) In at least one human pathogen, Pseudomonas aeruginosa, this apparently universal gram-negative “language” seems to be a means of regulating its virulence. (Tang, 37-43: Winson, 27-31) In fact bacteria may be able to communicate across species-for example, between Burkholderia cepacia and Pseudomonas aeruginosa. Exoproducts from the latter influence virulence-factor production by the former. If methods could be developed to interfere with quorum-sensing systems of gram-negative pathogens, a novel means of controlling their pathogenicity might be possible.
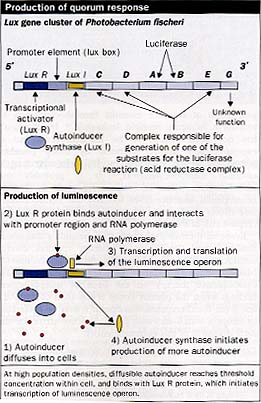
Understanding of this form of bacterial communication is based on a model used to describe the behavior of a marine organism, Photobacterium (Vibrio) fischeri, which expresses bacterial luciferase and thus luminesces when present in high concentrations in the light organs of certain fish. These organisms do not luminesce when the bacteria are present in the open water at low cell concentrations. Luminescence is achieved through the production of small diffusible compounds called autoinducers. (Kaplan, 1210-14) These autoinducers are N-acyl-L-homoserine lactone (NHL) molecules which, owing to their size, passively diffusible nature, and the properties of the gram-negative cell wall, are present in equal concentrations intracellularly and extracellularly. When the NHL-producing bacteria in the environment have reached a sufficiently high concentration, with the NHLsignal molecules exceeding a threshold concentrations (about 10 nmol/L) these autoinducers bring about the quorum response of bioluminescence through bacterial luciferase production. The autoinducer binds to an NHL-responsive transcriptional activator called LuxR, which in turn binds RNA polymerase and initiates transcription of the lux operon after binding to a promoter element. (Sitnikov, 801-12) The lux operon consists of several genes including LuxI-the autoinducer synthase. Thus after activation of the lux operon the basal levels of autoinducer rise substantially in a true positive feedback loop since the autoinducer synthase is also transcribed.
Earlier this decade LuxR homologues were found in Pseudomonas aeruginosa, and other bacterial species were shown to produce NHL molecules analogous to the autoinducer of P fischeri, with alterations in the acyl side-chain seeming to confer species specificity. (Swift, 511-20) Quorum-sensing systems have been identified in different bacteria, and their genes have been cloned and shown to have homology to both LuxR and LuxI of P fischeri. There are now more than a dozen homologues of LuxR and LuxI in the protein-sequence databases. LuxI homologues seem to synthesise NHLs with varying acyl side-chains, and the cognate LuxR homologues respond best to these particular autoinducers. Bacteria of medical importance known to produce NHL signals include: Pseudomonas aeruginosa, Yersinia enterocolitica, Burkholderia cepacia, Proteus mirabilis, Citrobacter freundii, Aeromonas hydrophila, Enterobacter agglomerans, Serratia liquefaciens, and Hafnia spp, with more being identified all the time.
What is controlled by quorum sensing in these bacteria has not been fully identified in every case, but Ps aeruginosa quorum sensing is used to control the expression of several extracellular virulence factors, including elastase, alkaline protease, haemolysin, cyanide, pyocyanin, and exotoxin A, with NHL signals also being used by Burkholderia cepacia to sense and respond to the density of Ps aeruginosa in the lungs of cystic fibrosis patients. (McKenny, 89-92) The rationale behind quorum-sensing systems in pathogenesis seems to be as follows: production of extracellular enzymes at low cell densities would be fruitless because the enzymes would diffuse away from the cells rapidly without converting much substrate in the environment into product useful for the bacteria. At high cell densities the concentration of extracellular enzyme or virulence factor would be high enough to bring about a change in the environment beneficial to the quorum of bacteria. In addition, the immunogenic exoenzymes and toxins are revealed to the immune system only once a sufficiently large number of bacteria have accumulated, in order to overwhelm host-defence mechanisms. That quorum sensing plays an important part in pathogenesis is borne out by the survival of neonatal mice colonised by Ps aeruginosa, which contained inactivating mutations in a gene homologous to LuxR, but succumbed to wild-type Ps aeruginosa infection. (Winson, 27-31)
The exciting aspect of quorum sensing is the realisation that virulent gram-negative organisms can potentially be rendered non-pathogenic by disruption of their quorum-sensing systems. On the basis of the model of quorum sensing in P fischeri, there are several theoretical ways in which this could be done. Autoinducer synthesis could be inhibited by inappropriate or synthetic NHL precursor analogues that would either have a higher affinity for the autoinducer synthase or be present in a sufficiently high concentration to block the production of the homoserine lactone molecules. Another, more attractive, strategy would be the prevention of activation of LuxR homologues by autoinducer homologues. As mentioned above, there seems to be an optimum NHL for a given LuxR-like protein, and any other homoserine lactone molecule brings about weaker activation or even inhibition of that particular quorum-sensing system. At least one such system already exists in nature. Researchers in Australia have discovered furanone compounds produced by the macroalga Delisea pulchra, which have structural similarity to NHL molecules and which interfere with the quorum-sensing system of Serratia liquefaciens. (Givskov, 18-22)
This it presumably does by binding to the S liquefaciens LuxR homologue, thereby preventing activation by the natural S liquefaciens NHLs. Yet another possible site of quorum-sensing disruption could be at the DNA-binding site of LuxR homologues, but this may prove to be difficult to target selectively because the DNA-binding region of LuxR has conformational similarity with several eukaryotic and prokaryotic transcription factors. The discovery of quorum-sensing systems in gram-negative bacteria not only has deepened understanding of the pathogenesis of infections caused by these organisms, but also may provide a means of treating these common and important infections.
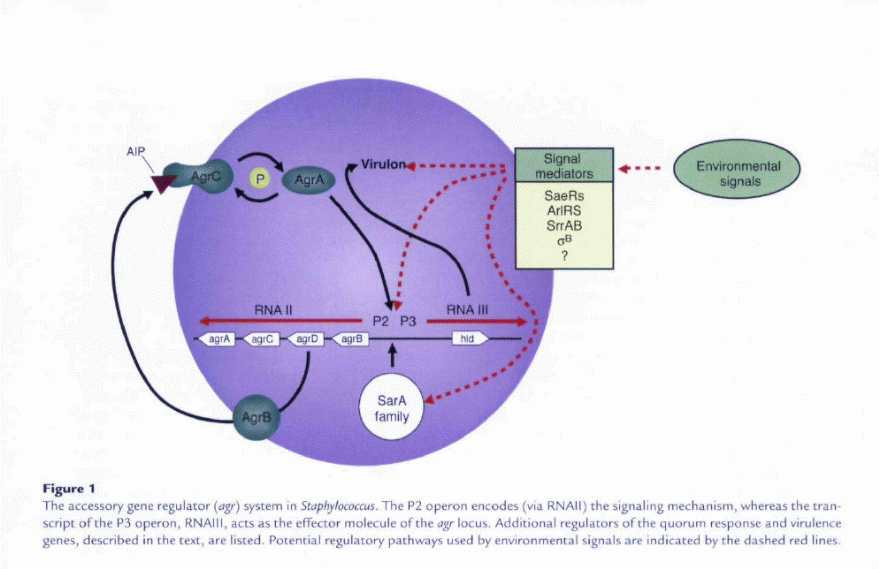
To improve their ability to cause this variety of human disease and to occupy numerous niches within the host, staphylococci have developed quorum-sensing systems that enable cell-to-cell communication and regulation of numerous colonization and virulence factors. (Raad, 1182-87) The staphylococcal accessory gene regulator quorum sensing system decreases the expression of several cell surface proteins and increases the expression of many secreted virulence factors in the transition from late exponential growth to stationary phase in vitro (Novick, 1429-49).
Expression was found to contribute to staphylococcal pathogenesis in several infection models, including murine subcutaneous abscesses (Bunce, 2636-40) and arthritis (Abdelinour, 3879-85), as well as rabbit endocarditic (Cheung, 1815-22). Expression of it also appears to be involved in the invasion and apoptosis of epithelial cells (Wesson, 5238-43). Interestingly, different groups, as defined by their production and recognition of distinct secreted signals, are associated predominantly with certain diseases (Novick, 1429-49). The reasons for this association between group and infection type are not yet clear, but a better understanding of this phenomenon may contribute to our understanding of the epidemiology of staphylococcal diseases. (Gocz, 1367-78)
Two primary transcripts, RNAII and RNAIII, are generated by the agr locus and originate from the P2 and promoters, respectively. The P2 operon encodes four proteins that generate the (sensing mechanism. AgrB is a transmembrane protein that appears to be involved m (a) processing of the agrD product into an octapeptide; (b) secretion ofthe autoinducing peptide (AIP) signal; and (c) modification of the AIP by the formation of a cyclic chiolactone bond between an internal cysteine and the carboxyl terminus AgrA and AgrC form a two-component regulatory system which the cransmembrane component, AgrC (histidine kinase), binds the extracellular AIP and in turn modulates the activity of AgrA, the response regulator.
Through an as-yet-undefined mechanism, AgrA activity then leads to greatly increased P2 and P3 transcription in the Iate-Iog phase of growth, when the concentration of the signai in the medium is high. Sequence variation in aB, aD. and agrC has led to the identification of at least four S. aureus agr specificity groups in whicb AIP produced by one group inhibits a^ expression in ocher groups (Novick, 1429-49). Increased transcription of the P3 operon results in dramatically increased levels of intracellular RNAIII. RNAIII encodes the toxin 6-hemolysin (via hid) buc, more importantly, increases the transcription (and in some cases, translation) of several secreted virulence factors, including TSS toxin-1 and other hemolysins.
However, some toxins, such as enterotoxins A and K, made typically in low concentrations and during exponential phase, are not regulated by RNAIII. Other secreted toxins, such as enterotoxins B, C, and D, are only partially upregulated by RNAIII and can be made in high concentrations independently of agr. At the same time, the expression of several cell surface virulence factors is decreased. It is easy to imagine the role that such coordination of virulence gene expression might play in certain infections, such as the formation of a walled-off furuncle. Initially, the staphylococci, present in small numbers, express their cell surface virulence factors in order to evade the host lminune system. For example, protein A binds the Fc portion of IgG, and clumping factor may help to form the walledoff infection site. When this site becomes depleted of nutrients due to increased bacterial numbers, the organisms increase secrered factor production, allowing the organisms to escape the walled-off site and spread through the host tissues.
The potential role of mediated quorum sensing is not as clear in other host environments. We and other researchers have shown through in vivo human and animal studies that ap- appears to be unnecessary in certain infections for the expression of secreted virulence factors (Yarwood, 1095-1101: Goerke, 1439-47). For example, expression in a rabbit abscess model was decreased at the same time that the animals developed TSS through exotoxin production (Yarwood, 1095-1101). In addition, mutant was just as effective at causing TSS as the isogenic wild-type organism. These studies do not rule out roles for agr in other aspects of these diseases, as will be discussed later in this review. But the studies do suggest that additional regulatory mechanisms are integral in regulation of both the quorum response and overall virulence of staphylococci. (Yarwood, 1095-1101: Goerke, 1439-47)
Additional Regulators of the Quorum Response
The quorum-sensing system has historically been assigned a central role in the model of S. aureus pathogenesis. Thus, studies of other known regulators of staphylococcal virulence have usually examined their interaction with the system. These additional regulators allow the organism to respond to environmental signals in addition to bacterial cell density, and sometimes counter activity. Among these regulators are the two-component systems (a family that includes AgrAC) chac allow scaphylococci CO sense and respond Co various environmental stimuli. SaeRS was the second two-component system involved in global regulation of virulence factors to be identified (Giraudo, 53-58) after AgrAC. sae mutants produce substantially less hemolysin and coagulase but have no effect on the production of RNAIII. However, the expression of at least one sae transcript was decreased in mucant, suggesting chat the sae system acts downstream of dgr (Giraudo, 246-50). It has been proposed, but not confirmed, chat the sae locus responds to several environmental stimuli, including high salt, low pH, glucose, and sub inhibitory concentrations of antibiotics (Giraudo, 15-22).
ArlRS comprises a third two-component system that appears co counter agr autoinduction by repressing production of hemolysins and exoenzymes (Fournier. 3955-64). Expression of arlRS was itself reduced in an agr mutant. An arlS mutant was enhanced for biofilm formation, despite the increased expression agr and the presumed downregulation of surface-associated adbesion factors. ArlRS also appears to regulate autolytic activity as well as the mulcidrug efflux pump NorA of: S. aureus. These data suggest that under certain conditions; agr activity may influence the resistance of S. aureus to antibiotics.( Fournier, 247-61)
A fourth two-component system, SrrAB, recently identified by our group (Yarwood, 1113-23) and others (Throup, 10392-401), was found to inhibit RNAIII expression and may itself be repressed hy agr. Mucancs of srrAB are unable to grow normally under anaerobic conditions (Throup, 10392-401), and expression of srrAB was shown to regulate genes involved in energy metabolism (Throup, 10392-401: Yarwood, 1113-23). The signal for the system may be menaquinone, an intermediate in the oxidative respiratory pathway. Thus, SrrAB may be one link between energy metabolism in the cell and the quorum response.
The second major family of regulators of staphylococcal virulence are the DNA-binding proteins, including stapbylococcal accessory regulator (SarA) and its homologs (Cheung, 1825-42). SarA, transcribed from three promoters within the same locus, was reported to be required for full agr transcription (Cheung, 3963-71). In several reports, SarA has been shown to affect the expression of a wide array of virulence genes, sometimes acting independently of agr to decrease the expression of several exoproteins. SarA is responsive to some environmental conditions through intermediate regulators (Deora, 6355-59) and likely affects expression of the locus accordingly.
Additional transcription regulators, whose interactions with the agr system deserve further investigation, are the repressor of toxins (Rot} and the alternative sigma factor B. Both of these have been shown co affect the expression of numerous virulence-associated genes. Rot appears co counter agr activity, and a mutation in raf was shown to partially restore the wild-type phenotype of mutant (McNamara, 3197-3203: Said-Salmi, 610-19). Sigma factor B, which responds to environmental stress (Chan, 6082-6089), also appears to at times counter agr activity in that it increases the expression of some exoproteins early in growth (Nicholas, 3667-69). Thus, the quorum response in staphylococci during infection occurs within the context of a complex regulatory network that continually modifies either agr activity itself or its downstream effects. Additional monitoring of gene expression and protein profiles in vivo will be required to understand this regulatory network that likely differs substantially from what has been described thus far in vitro.
Quorum sensing and staphylococcal bio-films
Many infections by staphylococci are not caused by the free-living organism but rather by groups of interacting cells termed bio-films. Bacterial bio-films are broadly defined as a community of ceils attached to either an abiotic or a biocic surface, are encased in a self-produced tnacrix, and generally exhibit an altered growth and gene expression profile compared with that of planktonic, or free-living, bacteria. Bio-film-associated infections have special clinical relevance, as they are generally resistant co antibiotic therapy and clearance by host defenses. In staphylococcal infections, these diseases include endocarditis (van Wamei, 73-79), osceomyelitis (Gillaspy, 3373-80), implanted device-related infections (Doniai, 167-93), and even some skin infections (Akiyama, 526-32).
Two stages of scaphylococcal biofilm formation have been described. The first stage involves actacbment ofeells to a surface. This stage of bio-film formation is likely to be mediated in part by cell wall-associated adbesins, including the microbial surface components recognizing adhesive matrix molecules (MSCRAMMs). The second stage of bio-film development includes cell multiplication and formation ofa mature, multi-layered, structured community. This stage is associated with production of extra-cellular factors, including the polysaccharide intercellular adhesion (PIA) component of the extra-cellular matrix. Detachment of cells from the established biofilm may then allow staphylococci to spread and colonize new sites.
One of the most intriguing areas of investigation is determining what impact quorum sensing has on the growth, development, and pathogenesis of scaphylococcal biofilms. There is mounting evidence that the phenotype and expression patterns may influence several aspects of biofilm behavior, including attachment of cells Co surfaces, biofilm dispersal, and even the chronic nature of many biofilm-associated infections. Indeed, many of the products involved in biofilm development, including C (-toxin, surface-associated adhesins, 5-hemolysin, and the autolysin AtlE (in S. epidermidis), are regulated by the system, at least in vitro. Furthermore, quorum sensing has been shown to be involved in biofilm development of several Gram-positive and Gram-negative bacteria, including Streptococcus mutans (Li. 2699-2708) and Pseudomonas aeru^nosa (Davies, 295-98).
The limited number of studies addressing the quorum response and sraphylococcal biofilms appear at first glance co be somewhat conflicting in their results and interpretation. Pratten et al. (Prarcen, 17) found little difference between wild-type S. aureus and an agr mutant in adherence to either uncoated or fibronectin-coated glass under flow conditions, even though hid was expressed. In another study, RNAIII expression decreased S. aureus adherence to fibrinogen under static conditions, but increased adherence to fibronectin and human endothelial cells in both static and flow conditions (Slienkman, 747-54). Vuong et al. (1688 1693) found that those S. aureus strains with a nonfunctional agr were much more likely to form biofilms under static conditions. A-toxin, positively regulated by the system, was recently shown to be required for biofilm formation under both static and flow conditions (O’Toole. 3214-17). In an experimental endocarditis study, RNAIII expression increased with increasing S. aureus densities in vegetations (Xiong, 668-77), confirming the cell density-dependent expression in vivo. Interestingly, expression of RNAIII also occurred through a mechanism independent of the AgrAC signaling system, suggesting that there are additional, unidentified in vivo signals chat regulate the quorum response. Taken together, these studies indicate that the precise role of agr expression in biofilm development is dependent upon the conditions m which the biofilm is grown, and suggest that differences in S. aureus strains may also introduce variability into the results.
Most infections of indwelling medical devices are caused by S. epidermidis, an organism with few exotoxins and for which the ability to form biofilms is considered the primary virulence factor (Raad, 182-87). Recently, Vuong et al. (706-18) found that disruption of the locus in S. epidermidis resulted in increased attachment of the bacteria to polystyrene, increased biofilm formation, and higher expression of AtlE, which enhances attachment to abiotic surfaces. They also confirmed chat the clinical isolate S. epidermidis 0-47, the strain of choice for studying biofilm formation in S. epidermidis, was an agr mutant. Interestingly, did not regulate PIA expression.
Even in conditions in which agr does not appear to contribute to biofilm growth or development, it may still affect the virulence of biofilm-associated bacteria. We have observed expression of the a^ system in conditions in which a^ did not appear to affect biofilm growth or structure (Yarwood, 39). It has been proposed that the production of 5-hemolysin, a molecule with surfactant properties that is encoded by the agr locus, may contribute to the detachment of cells from both 5. aureus and S. epidermidis biofilms (Vuong et al. 706-18). Should this in fact be the case, it has important clinical implications. Cells expressing an actively detaching from the biofilm not only may establish additional infection sites in the host but also may contribute Co the toxemia associated with acute staphylococcal infections, These cells are likely to express secreted virulence factors, including the toxins of S. aureus. On the other hand, cells chat remain in the biofilm and do not express agr may contribute to persistent, low-level infections, particularly in the case of S. epidermidis.
Evidence of selection for negative phenotype in chronic infections is emerging. Schwan et al. (Scbwan, 23-28) studied chronic wound infections using a murine abscess model. After establishing infections with a hemolytic (thus with a presumably functional agr locus) S. aureus strain, the number of nonhemolytic bacteria recovered from the wounds increased over time. The authors suggest that several of the nonhemolytic isolates had mutations in the locus, although this was not directly shown. In mixed-strain infection experiments using normal hyperhemolytic and nonhemolycic strains to inoculate the mouse, the population of hyperhemolytic isolates declined (44.0-9.3%) after 7 days, while the nonhemolytic group (presumably defective) increased (23.7-61.0%) over the same period of time. Conversely, in both this study and several previous ones, infections established with single strains resulted in decreased cell numbers recovered from the infection of the mutant compared with those of the wild-type strain.
Thus, in the mixed strain infection experiments, functions performed by the wild-type strain may assist the nonhemolytic group in establishing infection. Also consistent with a selection for a negative phenotype, expression of virulence factors in epidemic mechicillin-resistance strains is shifted away from extracellular toxins and enzymes coward expression of surface proteins and colonization factors (Lapakyriatou, 990-1000). Furthermore, mutants can f-frequently be found in isolates from clinical settings and arise spontaneously in micro (Somerville, 1430-37). When Vtiong et al. examined the correlation between a functional system and the ability of 5. aureus clinical isolates to adhere to polystyrene under static conditions, they found that only 6% of the isolates with a functional dgr system formed a biofilm in these conditions, compared with 78% of the defective isolates. Failure of the strains with functional loci to form a biofilm was thought to be due in part to the surfactant properties of the 6-hemolysin produced by these strains.
These studies are all consistent with the idea that whereas secreted virulence factors may be important during the acute phase of infection, loss of agr function may enhance the long-term survival of staphylococci in the host and contribute to persistent (often biofilm-associated) infections. The enhanced survival of agr mutants might be tied in part to the decreased production of immunostimulatory factors, such as superantigens, and increased expression of immune-evading factors, such as protein A. It has been proposed that use of inhibiting substances, such as AIP produced by staphylococci belonging CO a different agr group, might be beneficial in the treatment of acute staphylococcal infection (5). Such treatments may decrease the production of extra cellular virulence factors normally upregulated by agr expression. However, because loss agr activity normally correlates with increased expression of adhesion factors and decreased expression of potential dispersion factors, inhibition of the activity may instead result in the conversion of an acute infection into a chronic one. particularly in biofilm-associated infections.
Indeed, inhibition of activity increases attachment of both S, aurcHS and S. epidermtdis to polystyrene (36, 39) and enhances biofiim formation. Furthermore, in certain animal models of infection, expression of agr does not significantly affect the expression of virulence factors, as would be expected from in vitro data (10-12). Understanding these caveats, additional investigation into prohibition is still warranted, given that inhibition of agr activity, and thus extra cellular toxin and enzyme production, may be beneficial iii some acute infection types.
Taken as a whole, the literature suggests that the role of quorum sensing in staphylococcal infections may not always be immediately obvious, as it varies with infection type, host environment, and even time. For example, in models in which agr does not appear to strongly influence virulence, its expression (or lack thereof) may contribute to biofilm formation. Conversely, in models m which agr expression does not appear to affect biofilm formation, it may still regulate virulence factor production. This makes the development of relevant in vitro and in vivo models a challenging proposition. Yet it is critical that further investigation of this area take place. The expression of the agr quorum-sensing system, already known to affect virulence factor production, may also affect everything from antibiotic resistance to energy metabolism through its interaction with other staphylococcal gene regulators. Furthermore, the phenotype and expression patterns in both S. aureus and S. epidcnnidis biofilms may influence when these infections become chronic or acute. Future studies should include sampling of staphylococcal infections for phenotype, evaluation of expression patterns in vivo using animal models of infection, and further investigation of dysfunction in biofiim growth and development.
Works Cited
Abdelinour, A, Arvidson. S,, Bremell. T,, Ryden, C. and Tarkowski, A, 1993, The accessory gene regulator controls StaphylocoKUS aureiis virulence in a murine arthritis modd. Infect. Immun. 61:3879-3885.
Akiyama, H,, et al, 2003. Laser scanning microscopic observation of glycocalyx production by Stapbyiococcus aureus in skin lesions of builous impetigo, atopic dermatitis and pemphigus foliaceus. But.}. Dermatni.148:526-532.
Blevins.J,S,.et al, 2003, Roleof J JM in the f’a.iiiogenesa of Staphyiocixcus musculoskeletal infection. Infect. Immun. 71:516-523,
Bunce, C. Wlieeler, L.Reed, G.,MusserJ,,andBarg. N, l’J92, Murine model of coetaneous infection with gram-positive cocci, infect. Immun. 60:2636-2640
Chan, PR. Foster, S,J,. Ingham, E. and Clements, M,O, 1998, The Stapiniococcusaureus alternative sigma factor a” controls the environmental stress response bur not starvation survival or pathogenic>’ in a mouse abscess model,/ Bacteriol. 180:6082-6089.
Cheung, A, L, Bayer, M, (i.. and Heinrichs,J,H. 1997. genetic determinants necessary for transcription of RNAII and RNAlIi in the agr locus of Staphylococcuiaureus. J. Bactenol. 179:3963-3971.
Cheung, A,L, et al. 1994, Diminished virulence ofa.sar/agr mutant of StaphyiococcHSaureui in the rabbit model of endocardjti.s,/ Gin. invest. 94:1815-1822.
Cheung, A,L,, and Zhang. G, 2002, Global regulation of virulence determinants in Stapbyiococcus.lurcus by the SaiA protein tamik, front. BiMci. 7:1825-1842.
Davies, D,G,, et al, 1998. The involvement of cell-to-cell signals in the development of a bacterial biofiim. Science. 280:295 298.
Deora, R., Tseng, T, and Misra, T, 1997, Alternative transcription factor a”’^ fStaphyiiKoccus aureui characterization and role in transcription of the global regulatoi-y locus sar.J. Bacteriol. 179:6355-6359.
Doniaii, R, M, and Costerton J.W. 2002, Biofilms: survival mechanisms of clinically relevant microorganisms, CUn. Microbioi. Rev. 15:167-193,
Fournier, U, Klier, A,, and Rapoport, G, 2001, The two-component system ArlS-AriR is a regulator of virulence gene expression in Stapbytococcus aureus. Moi. Micmbiiil. 41:247 -261.
Fournier. B,, and Hooper. D,C 200(1. A new two-component regulatory system involved in adhesion, autolysis, and extracellular proceolytic AcxXwcy of Stapbyiococcusaureus.]. Bactenol. 182:3955-3964,
Fuqua WC, Winans SC, Greenberg EP. Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsiveness transcriptional regulators. J Bacteriol 1994; 176: 269-75.
Gillaspy, A,, et al, 1995, Role of the accessory gene regulator iagy] in pathogenesis of staphylococcal osteomyelitis. Infect. Immun. 63:3373-3380.
Giraudo, A,T,, Cal/.olari, A,, Caialdi, A,A,, Bogni, C. and Nagel. R. 1999, The Mf locus of Staphyiococcui aureus a two-component regulatory system, FEM,S Af/iroWo/. Lett, 177:15-22.
Giraudo, A,T,, Cheung, A,L,, and Nagel, R, 1997, The MP locus controls synthesis at the transcriptional level, Arci!. Mkrobioi. 168:53-58.
Giraudo, A,T., Mansilla, C. Than, A,, Raspaiiti, C, and Nagel, R, 2003. Studies on the expression of regulatory locus sae in Staphylococcus aureus. Curr. Microbioi. 46:246-250.
Givskov M, de Nys R, Manefield M, et al. Eukaryotic interference with homoserine lactone-mediated prokaryotic signalling. J Bacteriol 1996; 178: 6618-22.
Gocz, F, 2002, Staphylococcus and biofilms, Mol. Mkrobii’L 43; 1367-1378.
Goerke, C, Fliickiger, U,, Steinhuber, A,, Zimmerli. W, and Wolz, C, 2001, Impact of the regulatory loci agr. sarA and sae of Staphylococcus aureus on the induction of a-toxin during device-related infection resolved by direct quantitative transcript analysis. Moi Mtcrobiol. 40:1439 1447.
Lapakyriatou, H,, Va/,, [)., Simor. A,. Louie. M.. and McGavin. M.|. 2000. Molecular analysis of accessory gene regulator locus and balance of virulence factor expression in epidemic resistant Stdphtococcus aureus. J. infect. Dis. 181;990-1000.
LGoerke, C, et al, 2000, Direct quantitative transcript analysis of the region (Stapiiyiococcus aureus during human infection in comparison to the expression profile/” vttro. Infect, immun. 68:1304-1311.
Li. Y,H,, et al, 2002, A quorum-sensing signaling.system essential for genetic competence in Streptococcus mutans is involved in biofilm tormation,/ Bactenoi. 184:2699 2708.
McNamara. P,J,. Milligan-Monroe, K, Khalili,.S. and Proctor, R, A, 2000, Identification, cloning, and initial characterization of rot, a locus encoding a Regulator of virulence factor expression in Staphylococcus aiireus.J. Bactenoi. 182:3197-3203.
Nicholas, R.O.. et al. 1999, Isolation and characterization of a deletion mutant nfStaj’i’yiococtusaurfus. Infect. Immun. 67:3667-3669.
Novick, R,P, 2003. staphylococcal virulence. Mul. Mkmhiol. 48:1429-1449.
O’Toole. G.A. 2003, Alpha-toxin is required for biofiim Stapinii/coccus uureus. J. Unctenoi. 185:3214 3217.
Prarcen, J. Foster, SJ.. Chan. RF,, Wilson. M,, and Nair, S.P. 2m. Staphylococcus aureus accessory regulators; expression within biofilms and effect on adhesion. Micrabei infect. 3:63. 17.
Raad, I,, Alrahwan, A,, and Rolston, K, 1993, tapbyhicoKus cpidvrmidh: emerging resistance and need for alternative agents, Chn. infect. Pis. 26:1182 1187.
Said-Salmi, B, et al, 2003, Global regulation of Sfti/jfrv/ncoccHJiiun’ws genes by Rot. J-Baetenol. 185:610 619.
Scbwan, V, R, I.anghorne. H,D Stovtr, C.K. 2003. Loss of bemolysin expression in Staphyhcoccusaureus Jff- muianli correlatfs witb selective survival during mixed intVcrions in murine abscesses and wounds, FRMSbnniund. Med. Microbioi. 38;23-28.
Slienkman, B., ft al. 2002. (RNAIII) in Stjphylococcusaureus adherence under static and flow conditions./AJrt/, Microi’ioi. 51;747-754.
Somerville, G.A., et al, 2002. In vitro serial passage of Staphylociiccus jurcus: changes in pbysiolog)’, virulence’ factor production I84; 1430-1437.
Throup, J,P., et al. 2001. The srijSR gene pair from Staphyiococcus aureus: genomic and proteomic approaches to the identification and characterization of gene function. Biochemistry. 40:10392-10401.
van Wamei, W, et al, 2002, Regulation of type 5 capsular polysaccharides by agr and sarA in vitro and in an experimental endocarditic model, Microb. Patbog. 33:73-79.
Vtiong, C, Gorkc, C, Somcrvillc, G.A., Fischer. E.R., and Octo, M. 2(103, quorum sensing control of biofiim factors in Staphylococcus. J. Infect. DIS 188:706 718.
Vuiing, Q’.., Saonz, H.L Otro, M.2000. Impact of the quorum sensing system on adherence to polystyrene in Staphylococcus aureus. J. Infect. Dis. 182:1688 1693.
Wesson, C.A,, et al. 1998, Staphyiiicuaus aureus Agr global regulators influence internalization and induction of apoptosis. Infect. Immun. 66:5238-5243.
Xiong, Y,Q., et al. 2002. Activation and transcriptional interaction between RNAII and RNAIII in Stjpinlococcusaureus in vitro and in an experimental endocarditis model./ Infect. Dis. 186; 668-677.
Yarwood, J, M, and Schlievert, P,M, 2001, Identification of a novel tv’o-component Regulatory system that acts in global regulation of virulence factors of StuphyiocDccus Jiircus. J. Bactenoi. 183:1113 1123.
Yarwood, J.M,, McCormick, J,K,. Paustian. M,L,, Kapiir. V,. and Schlievcrt, P.M, 2002. Repression of the regulator/ Bacteriot. 184:1095-1101.
Yarwood.J, M, Bartels, D.J., Volper, E.M., Expression of the accessory gene regulator quorum sensing system in biofilms of Staphyiococcus aurvus. ASM Conference on Iliofiims2003. Victoria, British Columbia, Canada, 39.
Tang HB, DiMango E, Bryan R, et al. Contribution of specific Pseudomonas aeruginosa virulence factors to the pathogenesis of pneumonia in a neonatal mouse model of infection. Infect Immun 1996; 64: 37-43.
Winson MK, Camara M, Latifi A, et al. Multiple N-acyl-L-homoserine lactone signal molecules regulate production of virulence determinants and secondary metabolites in Pseudomonas aeruginosa. Proc Natl Acad Sci USA 1995; 92: 94, 27-31.
Kaplan HB, Greenberg EP. Diffusion of autoinducer is involved in regulation of the Vibrio fischeri luminescence system. J Bacteriol 1985; 163:1210-14.
Sitnikov DM, Schineller JB, Baldwin TO. Transcriptional regulation of bioluminescence genes from Vibrio fischeri. Mol Microbiol 1995; 17: 801-12.
Swift S, Winson MK, Chan PF, et al. A novel strategy for the isolation of luxI homologues: evidence for the widespread distribution of a LuxR:LuxI superfamily in enteric bacteria. Mol Microbiol 1993; 10: 511-20.
McKenny D, Brown KE, Allison DG. Influence of Pseudomonas aeruginosa exoproducts on virulence factor production in Burkholderia cepacia: evidence of interspecies communication. J Bacteriol 1995; 177: 69, 89-92.