Materials and methods
Animals
Two to three-year-old adult female common marmoset monkeys (Callithrix jacchus) with bodyweight ~350 g, and having a regular 28 d ovulatory cycles or gestation periods between 141 d and 145 d, were housed in cages for the study. Prior to do any experiment, permission from local and statutory Ethical Committee was obtained (check).
Ovaries were collected for analysis during the 5th and 10th d of the follicular phase (n=4) (Taylor et al. 2004), and at early (n=6), mid (n=3) and late (n=4) luteal phase (14-22 d) of the ovarian cycle (Wulff et al. 2001; Rowe et al. 2002). Additionally, ovaries from the pregnant animals were collected from the 2nd (n=5), 3rd (n=3) and 4th (n=6) week of pregnancy (Rowe et al. 2002). The ovary sections were prepared and processed for histological and immunohistological studies as shown earlier (Wulff et al. 2001; Rowe et al. 2002).
Treatments
To synchronize the timing of ovulation, marmosets were intramuscularly treated with 1 µg of a prostaglandin (PG) F2α analog, on days 12-15 of 20 d luteal phase to induce luteolysis. The day of the PG injection was designated as follicular day zero. A GnRH antagonist Antarelix, procured from Europeptides, Argenteuil, France, was dissolved in water to reach a concentration of 10 mg/ml. To provide a slow-release depot, Antarelix was administrated at a dose of 12 mg/kg body weight of animal by subcutaneously injecting on follicular day 0 (n=5) or day 5 (n= 5). Ovaries from the treated animals were collected 10 days following PG administration, corresponding to the periovulatory period in the control animals, which did not receive PG. Control marmosets were studied in parallel on day 0 (n=5), day ? and day 10 (n= 5) corresponding to PG administration.
All animals were injected i.v. with 20 mg BrdU (Roche Molecular Biochemicals, Essex, UK) prepared in saline. After 1 h they were killed with an i.v. injection of 400 µl of Euthetal (sodium pentobarbitone; Rhone Merieux, Harlow, Essex, UK). Ovaries were removed, weighed and immediately fixed in 4% neutral buffered formalin. After 24 h, the ovaries were transferred to 70% ethanol, dehydrated, and embedded in paraffin according to the standard procedures (include one reference here).
Immunohistochemistry
The effect of Antarelix treatment on the proliferative activity of the OSE was studied by quantifying the number of proliferating cells within the OSE area whose nuclei were stained with BrdU. For this section were prepared on glass slides as shown previously (Taylor et al. 2004; Wulff et al. 2001; Rowe et al. 2002).
For BrdU immunostaining, antigen retrieval was performed by pressure cooking using a Tefal Calypso pressure cooker (Tefal, Langley, Berks, UK). Sections in 0.01 M citrate buffer, pH 6, were kept for 6 min at a high-pressure setting (2). Slides were then left for 20 min in hot buffer and washed in Tris-buffered saline (TBS; 0.05 M Tris-HCl, pH? and 1.5 M NaCl). To reduce non-specific binding, sections were blocked in normal rabbit serum (NRS; 1:5 diluted with TBS containing freshly added 5% bovine serum albumin) for 30 min. Primary antibody, BrdU (mouse anti-BrdU; Roche Molecular Biochemicals, Essex, UK) was diluted 1:30 in TBS. Incubation was carried out overnight at 4 ºC. Slides were washed three times in TBS. Incubation with the secondary antibody rabbit anti-mouse IgG (1:60 diluted in NRS: TBS; DAKO Corp.) was carried out for 40 min at room temperature, followed by two items of washing in TBS. and incubation with alkaline phosphatase-anti-alkaline phosphatase complex (1:100 dilution in TBS; DAKO Crop.) for 40 min at room temperature. Slides were visualized using per slide, 500 µl nitro blue tetrazolium (NBT) solution containing 45 µl NBT substrate (Roche Molecular Biochemicals), 10 ml NBT buffer, 35 µl 5-bromo-3-chloro-3-indolyl-phosphate and 10 µl levamisole. Sections were counterstained with hematoxylin.
Quantification of Immunohistochemistry
To investigate proliferative changes of the OSE cells during the ovarian cycle and in response to GnRH-antagonist treatment, BrdU incorporation of the OSE was quantified. The proliferation of the OSE was detected all around the circumference and in all sections. At least 20 sections for each group of study were examined. The labeling index was determined by counting the number of OSE cells (labeled and unlabelled) in each section, with the 60X objective, and expressed as the percentage of labeled OSE cells. The proliferation index is expressed as a mean value for the number of OSE cells in each section.
Results
Effect of Antarelix treatment on OSE proliferation
Prior studies on Antarelix treatment in marmoset monkeys revealed significant reduction in BrdU labeling in ovarian endothelial cells and steroidogenesis in granulosa and theca cells of growing follicles (Taylor et al. 2004). It was a matter of interest to study how these changes would affect the overlaying OSE cells. A comparison of sections prepared from the ovaries of late follicular controls and the treatment groups revealed that the OSE cells of GnRH antagonist-treated ovaries from day 0-10 and from day 5-10 exhibited a decrease in the proliferation after both treatment schedules. Quantitative analysis confirmed significant decrease in cell proliferation in the OSE cells in both treated groups, and the effect was more pronounced in longer treatment schedule (day 0-10 treated group; P=0.001) (Figure)
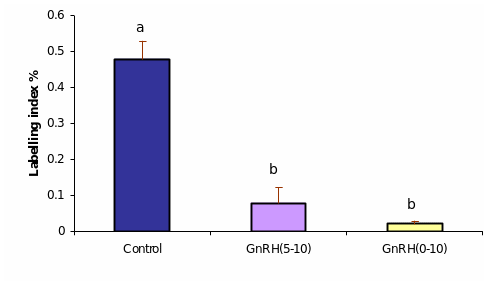
The data represent the mean % of total proliferation and the SEM from the mean. Different letters indicate a statistical differences between groups (P≤ 0.05).
OSE proliferation at different stages of the ovarian cycle and during pregnancy
OSE proliferation is influenced by the constantly changing underlying ovarian environment at different stages of follicular or luteal development, and during pregnancy. The proliferative index was calculated for the OSE cells during different follicular, luteal and early pregnancy phases. Results from this analysis revealed that the highest proliferation was obtained during the late follicular and early luteal phases. There was a gradual increase in the proliferation activity as the follicular phase advanced, reaching the maximum value towards the end of this phase (P≤0.05) (Figure A). On the other hand, a decreasing trend of proliferation was observed through the luteal phase, beginning at high rate of proliferation and thereafter decreasing in the proliferation index (P≤0.05) as the luteal phase approaches the end (Figure B).
To ascertain which underlying tissue is contributing to the OSE proliferation, the BrdU staining was examined at different regions of the OSE layer. During the follicular phase majority of the proliferating cells were detected over the large antral follicles which were very close to the surface (60-75μm), and a lower number of proliferating cells were found distributed randomly over the stroma. During the luteal phase most of the proliferating cells were located over the CL and little activity was observed over the stroma.
During all stages of pregnancy (2, 3 and 4 weeks), the OSE cells showed diminished or extremely low BrdU labeling, and correspondingly very low proliferation index as compared to the non-pregnant cycling animals (P≤ 0.05). The OSE proliferation was discernable mostly during the early pregnancy (Figure C). During early pregnancy a variation in the distribution of the proliferating cells within the OSE was also detected. The highest number of proliferating cells was found over the CL, whereas slightly lower number of evenly distributed cells was observed over stroma. At this pregnancy stage, large antral follicles were not detected close to the ovarian surface (Figure).

Different letters indicate statistical differences.
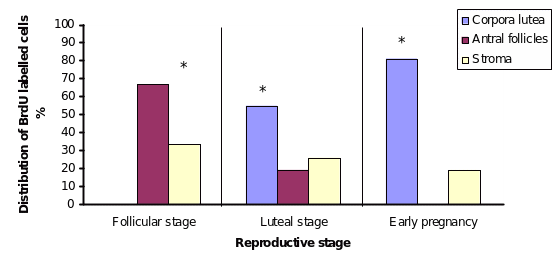
Distribution of BrdU labeling within the OSE cells at follicular and luteal phases and during pregnancy. Labeling distribution was investigated through different regions of the OSE layer, over corpora lutea, growing antral follicles and over ovarian stroma.
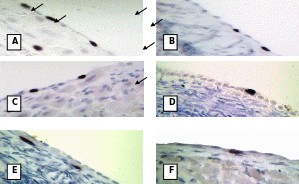
BrdU labeling within the OSE cells at follicular phase (A. over antral follicle and B. over stroma), Luteal phase (C. over mature CL and D. over stroma) and pregnancy (E. over stroma and F. over CL). Arrows indicate labeled cells (black stain). A, C, D and F at 400X; B and E at 200X.
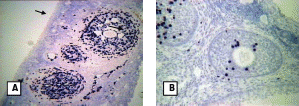
Histological section of ovary at A. follicular phase (arrow indicates high level of BrdU labeling in granulosa cells of antral follicle) and B. pregnancy (arrow indicates low level of BrdU labeling in granulosa cells of preantral follicle). A: at 40X and B: at 200X.
Morphological changes in the OSE cells
The distribution of proliferation markers in OSE cells considerably varied during different reproductive stages (follicular, luteal and pregnancy). The OSE morphology and its adhesiveness to underlying tissues are also expected to modify considering the extent of multiplication capabilities within a limited surface area in which the cells can expand. The microscopic observation revealed considerable variation in the shape of the OSE cells ranging from squamous to cuboidal. Moreover, the association between the OSE cells was affected by the ovarian cycle and the ovarian compartments. OSE cells over the large antral follicles which were located very close to surface were flattened in shape and disarrangement between these cells was noticed. There was no evidence of firm attachment with the underneath basement membrane (Figure A).Over the CL, OSE cells appeared as squamous with very weak association between the cells, and it was seen that attachment of these cells with the basement membrane was also loose (Figure B). The OSE cells observed in a region away from the follicular development and the ovarian events that are over the stroma, displayed highly zorganized topology of cuboidal shape, and attachment in between the cells and of the cells with underneath basement membrane was also firm (Figure C)

A. OSE covering large antral follicle (arrow indicates flattened shaped cells), B. over a mature CL (arrow indicates squamous shaped cells) and C. over stroma (arrow indicates cuboidal shaped cells). A and B: at 400X, and C: at 200X.